2ª Parte: Realidad o ficción: requerimientos de metionina para el camarón blanco del Pacífico Litopenaeus vannamei
Resumen
Se ha invertido un esfuerzo considerable en el desarrollo y validación de formulaciones de alimentos alternativos para camarones. Con base en la experiencia del IP con la transferencia de esta tecnología a los fabricantes de alimentos, la principal limitación es una comprensión deficiente o una falta de estudios definidos que identifiquen con precisión el requerimiento de metionina o aminoácidos azufrados totales (metionina + cisteína) del camarón. Muchos fabricantes de alimentos tienen el requerimiento de metionina establecido relativamente alto, lo que favorece la inclusión de harina de pescado costosa en lugar de fuentes de proteína no marina. Dadas las actuales preocupaciones económicas y sociales, este no es un enfoque sostenible. Hay varias publicaciones que evalúan los suplementos de metionina en los alimentos para camarones blancos del Pacífico. Sin embargo, estos artículos no brindan una definición clara del requerimiento, lo que es una limitación importante para que los fabricantes de alimentos acepten formulaciones de alimentos con bajo contenido de harina de pescado. Una teoría que se presenta a menudo es que los aminoácidos cristalinos se absorben y circulan en un patrón asincrónico con respecto a los de las proteínas intactas. Con base en la investigación actual, hemos demostrado que la absorción de aminoácidos en el camarón corresponde a la fisiología digestiva del animal. En el sentido de que los camarones son animales que se alimentan de forma semicontinua y procesan y digieren los alimentos muy rápidamente. Desde el inicio de la alimentación, se observó un aumento de los aminoácidos en la hemolinfa en 10 minutos, lo que confirma un procesamiento y digestión muy rápidos de los nutrientes. La depuración de los aminoácidos también fue relativamente rápida y los niveles de aminoácidos volvieron a los niveles de ayuno nocturno en 60 minutos posteriores al ayuno. Este ciclo de ciclado de nutrientes corresponde a los hábitos de alimentación semicontinua del camarón. Con base en los patrones de absorción y depuración de los aminoácidos, no hubo indicios de una absorción sincrónica de los aminoácidos suplementados; por lo tanto, los aminoácidos cristalinos deberían estar disponibles para el metabolismo. Por lo tanto, si podemos producir una dieta deficiente, deberíamos poder inducir una respuesta clásica a la dosis. En numerosos ensayos de crecimiento, publicados y no publicados, se ha evaluado la respuesta de los camarones juveniles a una variedad de fuentes de metionina. Estas fuentes incluyen Dl-metionina, metionina microencapsulada, metionina recubierta, péptidos sintéticos y diversas formas queladas. Sin embargo, en la literatura publicada hay poca consistencia en las respuestas o en estudios definitivos. Muchos de estos estudios presentan datos contradictorios o inconsistentes y, por lo tanto, no brindan evidencia sólida para un requerimiento definitivo. En nuestro laboratorio, en numerosos ensayos de crecimiento independientes, aumentar el nivel de metionina como suplemento o usar concentrado de proteína de maíz como fuente de proteína intacta, aumentar los niveles de metionina en la dieta produjo muy poca evidencia de mejora del crecimiento o ninguna consistencia en la respuesta. En la opinión de este autor, las respuestas inconsistentes se deben a otros factores y el requerimiento de metionina para esta especie es bastante bajo y puede no ser un aminoácido limitante en muchas formulaciones de alimentos comerciales. La hipótesis alternativa es que muchas de nuestras formas purificadas no están disponibles para los camarones o que las dietas y sistemas de prueba no son apropiados.
Evidentemente, existe la necesidad de comprender mejor el metabolismo de los aminoácidos en los camarones y definir los aminoácidos limitantes, así como las técnicas para definir de manera consistente los requerimientos de aminoácidos.
Palabras clave: metionina, requerimiento, camarones
Introducción
El camarón blanco del Pacífico, Litopenaeus vannamei, se considera la especie de camarón cultivado más importante del mundo y un producto de alto valor con una producción de alrededor de 4,5 millones de toneladas métricas en 2016 (Anderson 2016). Como se espera que el consumo de camarón siga aumentando, es vital desarrollar ingredientes alternativos sostenibles en las dietas para camarones a fin de respaldar la expansión de la industria del camarón (Achupallas et al. 2016b, Achupallas et al. 2016a). Desde un punto de vista nutricional, la popularidad del camarón blanco del Pacífico se debe a su adaptabilidad a una variedad de dietas, tolerancia a los alimentos de origen vegetal y capacidad para utilizar la productividad natural. Esta especie es muy tolerante a las dietas con una variedad de niveles de harina de pescado, incluidas las dietas sin harina de pescado (Morris et al. 2011, Roy et al. 2009, Samocha et al. 2004).
Mediante el uso de estrategias de reemplazo adecuadas (Davis 2007), varios estudios han demostrado que los niveles de harina de pescado y aceite marino se pueden reducir o eliminar de las dietas de engorde del camarón blanco del Pacífico. Actualmente, se puede utilizar una mezcla de proteínas vegetales más baratas y de alta calidad (por ejemplo, harina de soja extraída con solvente, proteína de soja especializada, concentrado de proteína de maíz, solubles de granos de destilería, harina de guisantes) o fuentes de proteína animal terrestre (por ejemplo, harina de subproductos avícolas, harina de carne y huesos, harina de sangre) para reemplazar con éxito la harina de pescado en los alimentos para camarones sin comprometer el crecimiento o la supervivencia (Sookying et al. 2013). Sin embargo, esto solo se puede lograr si se tienen en cuenta los cambios en los requisitos de nutrientes como los aminoácidos esenciales (en particular la metionina), el contenido energético de la dieta, los ácidos grasos esenciales y los minerales como el fósforo (Davis 2007). A pesar de la preponderancia de la información sobre formulaciones alternativas de alimentos (ver Sookying et al. (2013) para una revisión), aún falta una amplia aceptación de alimentos bajos en harina de pescado por parte de la industria de la acuicultura en general y la industria del camarón. Si bien los niveles de harina de pescado en las Américas se han reducido con el paso de los años, el nivel de harina de pescado en los alimentos para camarones en Asia es mucho más alto, es decir, el 20% o más de la dieta (observación personal). Es bien sabido que los suministros de harina de pescado no aumentarán, ya que la mayoría de las pesquerías están más allá de los límites sostenibles. Una parte de esta demanda se cubrirá con subproductos de la pesca que actualmente están subutilizados, así como con los productos de procesamiento de la acuicultura. Por lo tanto, si la acuicultura se va a expandir, la industria debe alejarse de la harina de pescado como fuente primaria de proteínas, particularmente en las dietas de producción.
Desde el punto de vista de la formulación de alimentos, uno debe preguntarse, ¿por qué nos encontramos en tal situación?
En primer lugar, hay muy pocos estudios de calidad que definan los requerimientos de aminoácidos esenciales para esta especie. El NRC informa un requerimiento de metionina dietética del 0,7% y un requerimiento de aminoácidos azufrados totales (TSAA, metionina + cisteína) del 1% tanto para el camarón Kuruma como para el camarón tigre, pero no informa uno para el camarón blanco del Pacífico (NRC 2011). Fox et al. (2011) informaron sobre una serie de estudios diseñados para evaluar la digestibilidad, la lixiviación y el requerimiento de metionina del camarón blanco del Pacífico. Aunque esta investigación confirmó la digestibilidad de una variedad de suplementos de metionina, no pudieron demostrar una deficiencia de metionina. Forster y Dominy (2006) evaluaron tres fuentes de metionina suplementadas a una dieta basal. Si bien informaron una deficiencia en la dieta basal que contenía 0,45% de metionina. Esta afirmación se basó en un contraste en el que se agruparon todos los datos suplementarios. Sin embargo, los autores también informan que los resultados de ANOVA fueron > 0,1 para los datos de crecimiento informados. Más recientemente, Façanha et al. (2016) informaron sobre los resultados de un ensayo de metionina en tres densidades en tanques al aire libre. No observaron respuesta a la dosis en dos de las densidades y un aumento "lineal" en la ganancia de peso en la tercera, con el requerimiento estimado en 0,72 (1,19 % TSAA) a 0,81 (1,28 TSAA) utilizando una dieta de 36 % de proteína. En opinión de este autor, esto es bastante extraño ya que la respuesta no debería verse afectada por la densidad y no hubo una meseta de rendimiento clara en el camarón para el que se observó una respuesta. En otra publicación reciente, Lin et al. (2015) informaron sobre tres experimentos con tres tamaños diferentes de camarones. No encontraron casi ninguna diferencia en la ganancia de peso en todos los experimentos y tratamientos. Sin embargo, utilizando un análisis de regresión, presumiblemente de los valores medios, se informó una "respuesta a la dosis" de metionina que estimó un requerimiento de metionina del 0,91 % de la dieta o del 2,28 % de la proteína en una dieta con 0,5 % de cisteína. Una vez más, se trata de datos muy débiles, que pueden no haber sido analizados utilizando los métodos más apropiados (es decir, regresión sobre las medias de los tratamientos), la estimación del requerimiento es mucho mayor que la informada para otras especies y los resultados no fueron repetibles en todas las clases de tamaño.
La definición de un requerimiento se complica aún más por las publicaciones de revistas especializadas que han publicado una variedad de artículos que pueden ser malinterpretados. Por ejemplo, un artículo reciente presenta un estudio de los niveles de aminoácidos esenciales (EAA) en alimentos comerciales. Los autores indicaron que el nivel promedio de TSAA encontrado en alimentos para camarones blancos del Pacífico fue de 3,36 % de la proteína. Además, respaldaron una recomendación de niveles extremadamente altos de metionina y otros EAA. Esto alcanza un nivel tan alto de metionina y TSA. Sin embargo, estos números claramente entran en conflicto con los niveles de cuerpo entero, el valor de reemplazo de cisteína, así como los niveles publicados para otras especies.
Por lo tanto, es muy probable que estos números tergiversen los requerimientos, y dichos informes no le hacen ningún bien a la industria. Con la excepción de la lisina (Fox et al. 1995), hay pocos estudios clásicos de requerimientos que refuten estas afirmaciones. Sugerimos que se debe iniciar un esfuerzo concertado y una evaluación sistemática y racional de la limitación de AA en un esfuerzo por definir el requerimiento en el camarón blanco del Pacífico a través de un enfoque clásico. Estos datos permitirían a los formuladores de alimentos tener la confianza para intercambiar fuentes de proteínas en función de los perfiles de AA y el costo.
Con el apoyo de una variedad de fuentes de financiación, mi laboratorio ha desarrollado sistemáticamente formulaciones de alimentos a base de soja para camarones (Sookying et al. 2013). En numerosos ensayos con fuentes de proteína intactas, no hemos demostrado ninguna respuesta a las sustituciones, que han llevado los niveles de metionina y aminoácidos azufrados totales a niveles muy bajos, sin una disminución en el crecimiento o la supervivencia. Recientemente, nuestro laboratorio ha dedicado un esfuerzo considerable a buscar aminoácidos limitantes en los alimentos para camarones. A continuación, se resumen datos experimentales recientes que intentan dilucidar el requerimiento de metionina y evaluar la eficacia de los aminoácidos cristalinos.
Métodos
Preparación de la dieta
Las dietas basales utilizadas en los diversos ensayos se diseñaron para contener 34% de proteína y 9% de lípidos utilizando principalmente harina de soja y harina de pescado o harina de subproductos avícolas como fuentes de proteína primarias (Tabla 1, 2, 3 y 4). Con excepción de la metionina, las dietas basales y de prueba se formularon para satisfacer los requerimientos nutricionales del camarón blanco del Pacífico. Las dietas de prueba se prepararon en el laboratorio de alimentos de la Universidad de Auburn, Auburn, AL, EE. UU., utilizando prácticas estándar. En resumen, los ingredientes secos previamente molidos y el aceite se mezclaron en una mezcladora de alimentos (Hobart Corporation, Troy, OH, EE. UU.) durante 15 minutos. Luego se incorporó agua caliente a la mezcla para lograr una consistencia adecuada para la granulación. Cada dieta se granuló a presión utilizando una picadora de carne y una matriz de 3 mm. Después de la granulación, las dietas se secaron hasta un contenido de humedad del 8-10% y se almacenaron a 4 C.
Sistema experimental
En todos los ensayos, se colocaron camarones blancos del Pacífico en tanques de 75 L que formaban parte de un sistema de recirculación interior de 2500 L (depósito, filtro de perlas, filtro biológico fluidizado y bombas de circulación y calentador). Cada dieta se ofreció a los camarones en tres a cinco acuarios replicados durante la duración del ensayo. La temperatura del agua se mantuvo alrededor de los 28 C utilizando un calentador sumergido de 3600 W (Aquatic Eco-Systems Inc., Apopka, Florida, EE. UU.). El oxígeno disuelto se mantuvo cerca de la saturación utilizando piedras difusoras en cada acuario y el tanque de sumidero utilizando una línea de aire común conectada a un soplador regenerativo. El oxígeno disuelto y la temperatura del agua se midieron dos veces al día utilizando un medidor de oxígeno/temperatura digital YSI-55 (YSI corporation, Yellow Springs, Ohio, EE. UU.) mientras que el pH, el TAN y el nitrito-N se midieron una vez por semana.
Aminoácidos libres (AA) de la hemolinfa
Para determinar los niveles de aminoácidos libres (AA) en la hemolinfa de los camarones, se utilizaron grupos replicados de cuatro camarones para cada punto de recolección. Los camarones en un punto de tiempo determinado se capturaron con una red y se colocaron en una suspensión de hielo para anestesiarlos, después de lo cual se recolectó hemolinfa de los segmentos abdominales utilizando una jeringa y una aguja de calibre 25.
Para este trabajo, se cargó previamente una jeringa de 1 ml con 0,5 ml de un anticoagulante y se utilizó para recolectar aproximadamente 0,5 ml de hemolinfa. Para mantener los datos semicuantitativos, el ensayo uno registró el volumen, mientras que en los ensayos siguientes, el peso del anticoagulante y la hemolinfa se determinaron gravimétricamente. La solución anticoagulante (Liu et al 2004) contiene 30 mM de citrato de sodio tribásico dihidrato (Sigma S4641); 0,34 M de cloruro de sodio (NaCl); 10 mM de EDTA (ácido etilendiaminotetraacético) (Sigma, E9884) en agua desionizada (DI). Para prepararlo, se pesan 4,4115 g de citrato de sodio tribásico deshidratado, 9,945 g de cloruro de sodio, 1,461 g de EDTA y se disuelven en 500 ml de agua DI. Para obtener muestras de aminoácidos libres de la hemolinfa, se centrifugó y decantó la hemolinfa recolectada. Se agruparon las muestras de sangre de los camarones de un solo acuario, lo que dio como resultado 2 o 3 muestras por tratamiento, cada una con sangre de 3 o 4 camarones.
Se realizaron tres ensayos con la dieta basal (BD) y la BD con AA seleccionados. Para cada ensayo, los camarones fueron preacondicionados a las dietas durante cuatro días. La noche anterior, se limpiaron los tanques para eliminar cualquier fuente de alimento. Al día siguiente y antes de alimentar a un grupo de camarones, se les extrajo sangre para determinar los niveles de AA libres en ayunas, se les ofreció alimento a los camarones restantes durante períodos de tiempo predeterminados y luego se les retiró el alimento y se los dejó en ayunas.
Ensayos de crecimiento
Se llevó a cabo una serie de ensayos de crecimiento utilizando camarones juveniles. Se ofrecieron dietas utilizando un protocolo de alimentación estándar con 4 comidas por día. Las dietas se ofrecieron a los camarones en un nivel adecuado según el rendimiento histórico, así como el consumo observado del alimento. Al concluir los ensayos de crecimiento, los camarones se pesaron, enumeraron y congelaron para su posterior análisis.
Análisis bioquímico
Las muestras de hemolinfa y camarones se congelaron hasta que se necesitaron. Las muestras de tejido que requerían análisis se enviaron al laboratorio de la Universidad de Missouri.
Análisis estadístico
Todos los datos se analizaron utilizando SAS (V9.4. SAS Institute, Cary, NC, EE. UU.). Los datos de los ensayos de crecimiento se analizaron utilizando un análisis de varianza unidireccional para determinar diferencias significativas (P < 0,05) entre las medias de los tratamientos, seguido de la prueba de rango múltiple de Student-Neuman-Keuls para distinguir diferencias significativas entre las medias de los tratamientos.
Resultados y discusión
Debido a la limitación de los datos disponibles sobre los requerimientos de metionina de los camarones, el trabajo inicial se concentró en confirmar los niveles de aminoácidos en la hemolinfa de los camarones. Se presenta un pequeño componente de estos datos que demuestra los aumentos después de la alimentación y la eliminación de aminoácidos una vez que los camarones estuvieron en ayunas (Figuras 1, 2 y 3). Hemos evaluado tanto a largo plazo (hasta 4,5 horas después de la alimentación) como a corto plazo (mientras se alimentan).
En el primer ensayo presentado en este resumen, los camarones ayunaron durante la noche para permitir la identificación de los niveles basales de AA. Después del ayuno nocturno, se tomaron muestras de los camarones (-30 minutos), luego se les proporcionó alimento durante 30 minutos, después de lo cual se tomó otra muestra (0 minutos) y se determinaron los niveles de AA posteriores a la alimentación (0 a 270 minutos). Como la mayoría de los AA siguieron un patrón similar, la suma de AA en la hemolinfa se presenta en la Figura 1. Está claro que todos los AA estaban cerca de su pico después de la alimentación y el nivel disminuyó en 60 minutos hasta casi los niveles de ayuno. Sin embargo, los niveles de AA se monitorearon durante 270 minutos para los cuales no hubo picos secundarios para la metionina o los otros aminoácidos suplementados (lisina, arginina o taurina).
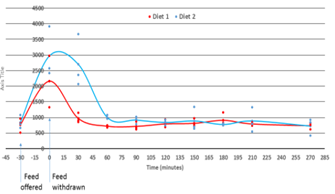
Figura 1. Se ofreció una ración a los camarones (7 g) durante un período de 30 minutos después de ayunar durante la noche. Se presentan los niveles totales de AA (µg/ml) de hemolinfa para las dietas 1 (basal) y 2 (basal + lisina, metanfetamina, arginina y taurina)
Para definir mejor el proceso de absorción y evaluar si puede haber un pico asincrónico durante la absorción, se realizó un segundo ensayo, que simplemente observó la absorción de AA durante un período de 1 hora (Figura 2). Nuevamente, los camarones ayunaron durante la noche y se tomó una muestra de su hemolinfa para determinar los niveles basales (-60 minutos). Luego, se les ofreció alimento a los camarones durante un total de 60 minutos durante los cuales se tomaron muestras intermitentes. Estos datos se presentan para la metionina (Figura 2), aunque la tendencia fue similar para todos los aminoácidos. Se siguieron tanto la dieta basal suplementada (Dieta 1) como la dieta suplementada con DLmetionina (Dieta 3). Aunque el nivel de AA es consistentemente más alto en la dieta suplementada, el patrón de absorción de AA es similar con un claro aumento en los niveles de AA en 10-15 minutos después del inicio de la alimentación. Esto es bastante diferente de lo que ocurre en las especies de peces, en las que la digestión y la absorción se retrasan varias horas, principalmente debido a la retención de alimentos en el estómago.

Figura 2. Se ofreció una ración a los camarones (30,6 g) después del ayuno durante la noche. Se presentan los niveles de metionina (µg/ml) de hemolinfa durante un período de 60 minutos cuando la orina está disponible para los camarones para las dietas 1 (basal) y 3 (basal) + 0,1 mg de metionina.

Figura 3. Niveles de metionina (µg/ml) de hemolinfa de camarones (16,6 g) alimentados con las Dietas 1 (Basal) y Dieta 3 (Basal + DL metionina). Se tomaron muestras de los camarones después de un ayuno nocturno (T-15) y se les ofreció alimento durante 15 minutos (T-15 a TO) y luego ayunaron durante un período de 60 minutos.
Los datos presentados demuestran claramente que la absorción de AA en el camarón es mucho más rápida que en las especies de peces, ya que aparece en la hemolinfa en un plazo de 10 a 15 minutos. Esto se debe principalmente al procesamiento rápido y continuo de los alimentos por parte del camarón. Según los resultados de estos estudios, no hay evidencia de que el camarón no pueda utilizar AA libre o de que se absorba de forma asincrónica.
Actualmente, hay pocos estudios publicados sobre el requerimiento de metionina del camarón blanco del Pacífico. Los pocos estudios que tratan sobre suplementos de metionina no han logrado establecer lo que se consideraría un requerimiento de metionina fuerte y convincente para esta especie.
Por lo tanto, el segundo componente de la investigación se diseñó para evaluar la eficacia de varias fuentes de metionina, así como para intentar identificar un requerimiento dietético. La Tabla 1a presenta una serie de dietas que se complementaron con una variedad de fuentes de metionina, incluidas: DL-metionina, metionina recubierta y una dieta modificada que utiliza proteínas intactas. La composición proximal y de AA de las dietas de prueba se presenta en la Tabla 1b, con la dieta basal analizada en 0,49% de metionina (0,41 cisteína) y las dietas restantes en 0,64 a 0,68% de metionina. Estas dietas se ofrecieron a camarones de 0,5 g durante una prueba de crecimiento de 42 días, después de la cual se evaluó el perfil de AA del músculo de la cola de los camarones y el hepatopáncreas. El crecimiento y la supervivencia fueron muy buenos, con un aumento de peso de más del 1000% observado con un buen FCR y supervivencia (Tabla 1c). Sin embargo, no se observaron diferencias en las tasas de crecimiento, alimentación o supervivencia. Para determinar si hubo cambios en la composición tisular, se determinaron los niveles de AA del músculo del camarón y el hepatopáncreas. No se observaron diferencias ni tendencias en los niveles de metionina de estos tejidos. Estos resultados indicaron que no hubo diferencias en la deposición en todos los tratamientos.
Tabla 1a. Composición (g/100 g tal cual) de las dietas de prueba.

Tabla 1b. Análisis de las dietas de prueba.

Tabla 1c. Respuesta de los camarones juveniles (0,55 + 0,04 g) a varias dietas de prueba durante un ensayo de crecimiento de 42 días. Medias de tres grupos de camarones replicados.

Para determinar si podíamos inducir una respuesta a niveles graduados de metionina, el primer ensayo de respuesta a la dosis utilizó tanto metionina DL como metionina microencapsulada (Tabla 2a). Este ensayo se repitió dos veces y se informaron los datos del segundo ensayo. El análisis de la composición proximal y de AA de las dietas de prueba se presenta en la Tabla 2b. Los niveles de metionina oscilaron entre 0,54 % (0,42 % de cisteína) en la dieta basal hasta 0,83 % de metionina para la dieta suplementada con metionina DL y 0,81 % de metionina para la dieta con metionina microencapsulada. La dieta 10 se diseñó como una dieta de proteína intacta que utilizaba concentrado de proteína de maíz que aumentó la metionina al 0,62 % de la dieta.
Al concluir un ensayo de crecimiento de 42 días (Tabla 2c), el camarón creció de 0,21 g a un peso máximo de 6,26 g. La supervivencia y el crecimiento fueron muy buenos en este ensayo y hubo un gran aumento en la ganancia de peso o el reemplazo de tejido (2467 a 2832%). Sin embargo, la respuesta no fue típica de un estudio de respuesta a la dosis. Es decir, no hubo un aumento en la ganancia de peso correspondiente a los aumentos en los niveles de metionina. La composición proximal y el perfil de AA de todo el camarón también se evaluaron para este ensayo con una ligera disminución en el nivel de proteína en el camarón y diferencias significativas en los niveles de metionina de todo el camarón. En este ensayo, la dieta basal resultó en una depresión significativa en los niveles de tejido corporal total que fue menor que la mayoría de las otras dietas. Según el análisis de regresión, hay un aumento general a medida que el nivel de metionina aumenta desde el 0,54% de la dieta, aunque no una respuesta muy fuerte (R2 = 0,54). Esto puede indicar una ligera deficiencia de la dieta basal.
Tabla 2a. Composición (g/100 g tal cual) de las dietas de prueba

Tabla 2b. Composición proximal y de aminoácidos (AA) de las dietas de prueba (g/100 g tal cual)

Tabla 2c. Respuesta de los camarones juveniles (0,21 + 0,017 g) a varias dietas de prueba durante un ensayo de crecimiento de 42 días. Medias de cuatro grupos de camarones replicados. Estas dietas se evaluaron en dos ensayos, se presentan los resultados del segundo ensayo. Las medias dentro de la misma columna con letras diferentes son significativamente diferentes según la prueba de Student-Newman-Kuels. La prueba T de Dunnett no indicó que ninguno de los valores dentro de una columna fuera diferente del de la dieta basal.

En el último ensayo de crecimiento, la dieta basal (Tabla 3a) fue similar a la utilizada en otros ensayos y se esperaba que tuviera un nivel similar de metionina. La primera serie de dietas se formuló con niveles crecientes de concentrado de proteína de maíz (0, 5, 10, 15, 20 y 25%) como sustituto de la harina de soja, lo que permitió un aumento gradual de la metionina en la dieta. En el segundo conjunto de dietas, la dieta basal se complementó con altos niveles de DL-metionina (0, 0,1, 0,2, 0,4, 0,6%).
Estas dietas se ofrecieron luego a camarones juveniles (0,85 g) durante un ensayo de crecimiento de 42 días. En este ensayo, los camarones a los que se les ofreció las dietas suplementadas con DL-metionina y el nivel más alto de concentrado de proteína de maíz fueron significativamente más pequeños que los camarones a los que se les ofreció la dieta basal. Nuevamente, no hubo una tendencia en los datos en términos de un mejor crecimiento debido a los suplementos de metionina, ya sea de proteína intacta o de DL-metionina. Una vez más no hay consistencia en los datos, que apuntan a que la dieta basal no es deficiente o, en el mejor de los casos, marginalmente deficiente.
Tabla 3a. Composición (g/100 g tal cual) de las dietas de prueba

Tabla 3b Respuesta de los camarones juveniles (0,85 g) a varias dietas de prueba durante un ensayo de crecimiento de 42 días. Se presentan las medias de cuatro grupos de camarones replicados. Las medias con diferentes superíndices dentro de una columna son significativamente diferentes según la prueba de Student-Newman-Kuels. Las medias con un * indican una diferencia significativa en la respuesta en comparación con la de la dieta basal.

Conclusión
Las investigaciones realizadas hasta la fecha en nuestro laboratorio han demostrado que la absorción de aminoácidos en el camarón corresponde a la fisiología digestiva del animal, ya que los camarones son alimentadores semicontinuos que procesan y digieren los alimentos muy rápidamente. Desde el inicio de la alimentación, se observó un aumento de los aminoácidos en la hemolinfa. La depuración de aminoácidos también fue relativamente rápida y los niveles de aminoácidos volvieron a los niveles de ayuno nocturno en 60 minutos. Este ciclo de nutrientes corresponde a los hábitos de alimentación semicontinua del camarón. Con base en los patrones de absorción y depuración de los aminoácidos, no hubo indicios de una absorción sincrónica de aminoácidos suplementados.
Los resultados de una serie de ensayos de crecimiento realizados con una variedad de suplementos, así como fuentes de proteína intacta, produjeron una variedad de niveles de metionina dietética, pero no se obtuvieron datos consistentes y persuasivos. En cuatro ensayos de crecimiento independientes, el aumento del nivel de metionina como suplemento puro o el uso de concentrado de proteína de maíz como fuente de proteína intacta, produjo muy poca evidencia de mejora del crecimiento. Claramente, parte del problema es la variación inherente a los ensayos con camarones. Esto lleva a tres posibles conclusiones: 1) la dieta basal es abundante o solo marginalmente deficiente en metionina y TSAA, lo que hace que la suplementación con metionina sea ineficaz y la identificación de una deficiencia más problemática, 2) otro aminoácido es el primero en limitar, por lo que también hace que la suplementación con metionina sea ineficaz, o 3) la metionina cristalina no funciona en los alimentos para camarones, aunque esto se contradice con el trabajo realizado con lisina en el que tanto la lisina cristalina como la intacta produjeron una respuesta dietética. Dado el número de intentos, así como los informes en la literatura de respuestas deficientes a los niveles de metionina, solo se puede concluir que la deficiencia es igual o inferior a la de nuestras dietas basales (0,54% de metionina y 0,42% de cisteína). Estos datos no corresponden a los informes en la literatura que indican un mayor requerimiento de metionina. En opinión de este autor, necesitamos un método y una dieta que induzcan una deficiencia de manera constante y repetida. Hasta que esto no se desarrolle no habrá acuerdo sobre un requerimiento dietético.
Lamentablemente, a menos que podamos demostrar una respuesta a la dosis de metionina (u otros AA), los formuladores de alimentos seguirán formulando dietas en exceso con harina de pescado y limitarán el uso de harina de soja. Por lo tanto, proponemos dar un paso atrás y desarrollar una dieta de prueba con aminoácidos cristalinos para evaluar la clasificación de los aminoácidos limitantes. Este enfoque proporcionará la base para una evaluación sistemática de los requisitos de EAA para el camarón blanco del Pacífico.