13ª Parte: Un meta análisis de los requisitos de aminoácidos esenciales del pescado
Resumen
Existen amplias variaciones en las estimaciones publicadas de los requisitos de aminoácidos esenciales (EAA). Se cree que las variaciones se originan a partir de diferentes elecciones en el modo de expresión, la variable de respuesta y el modelo matemático. Aquí llevamos a cabo un metaanálisis de los ensayos de dosis-respuesta basados en el crecimiento de 10 EAA en 22 especies de teleósteos: se revisaron 249 estudios. Los datos publicados se ingresaron en una hoja de cálculo y se volvieron a calcular de manera estándar y sistemática para permitir las comparaciones. Las unidades de requerimiento consideradas fueron el porcentaje de dieta seca, g de EAA por MJ de energía digestible (DE) y g de EAA ingeridos por kg de peso corporal metabólico (MBW) por día. Las variables de respuesta incluyeron el crecimiento en g por kg MBW por día y el coeficiente de crecimiento de la unidad térmica (TGC). También se compararon cuatro modelos matemáticos: modelo de cal rota (BLM), modelo cuadrático (QM), modelo cuadrático roto (BQM) y modelo cinético de saturación (SKM). Los resultados primero indican diferencias importantes en la calidad del estudio, ya que el 54 % de los artículos revisados se excluyeron del metanálisis, muchas veces debido a un crecimiento deficiente o falta de información.

Además, el conjunto de datos final estaba muy fragmentado: el 31 % de los estudios se refería a la trucha arcoíris, y la lisina era el foco del 29 % de todos los estudios, lo que dejaba algunas especies y EAA poco cubiertos. Las comparaciones de las estimaciones de requisitos muestran variaciones importantes entre estudios, incluso dentro de las especies. Con tal variabilidad no hubo diferencia en las estimaciones de requisitos calculadas con diferentes variables de respuesta. De igual forma, esta variabilidad no fue diferente entre los tres modos de expresión, ni tampoco entre modelos matemáticos. Sin embargo, hubo efectos significativos del diseño experimental en la calidad de ajuste de los modelos. Específicamente, los experimentos que no lograron producir una curva dosis-respuesta clara y estancada tenían una probabilidad mucho mayor de producir resultados absurdos (por ejemplo, requerimiento negativo). Finalmente, el presente estudio enfatiza la necesidad crítica de un sistema global, estándar y sistemático para informar y capturar los resultados de los ensayos de nutrición.
Palabras clave: Metanálisis, aminoácido, requerimiento.
1. Introducción
Las formulaciones de alimentos acuícolas se refinan, actualizan y ajustan constantemente para reflejar el progreso en la ciencia de la nutrición, así como la variabilidad en el precio y la disponibilidad de los ingredientes. Los alimentos deben seguir siendo rentables y entregar nutrientes de manera eficiente al animal para promover el crecimiento y la salud al tiempo que minimizan la descarga de nutrientes en el efluente. Como resultado, los alimentos para peces se formulan cada vez más a nivel de nutrientes, como se hace comúnmente en otros animales, por ejemplo, las aves de corral. Naturalmente, el primer requisito previo para este objetivo es un conocimiento preciso de los requisitos de aminoácidos esenciales (EAA).
Sin embargo, varias revisiones identificaron brechas significativas en nuestra comprensión de los requisitos de EAA de los teleósteos y destacaron la gran variabilidad en las estimaciones de los requisitos de EAA entre y dentro de las especies (Bureau y Encarnação, 2006; Cowey, 1994; Lall y Anderson, 2005; NRC , 1993, 2011; Wilson, 1989). Por ejemplo, en la trucha arcoíris Oncorhynchus mykiss, el requerimiento de lisina varía de 1,3 a 2,9 % de la dieta (3,7 a 7,3 % de proteína cruda, PC), y las estimaciones de requerimiento de arginina van de 1,4 a 2,2 % de la dieta.
Es probable que las variaciones genéticas solo representen una fracción de tales estimaciones diferentes. Más bien, las razones que explican estas discrepancias se pueden encontrar en diferentes enfoques metodológicos. El cálculo de una estimación de requisitos requiere elegir un modo de expresión, una variable de respuesta y un modelo matemático para describir su relación. Los modos de expresión incluyen porcentaje de la dieta o proteína cruda, o en g/MJ de energía digestible. La elección del modo de expresión es un tema de mucho debate y refleja los diversos supuestos que hacen los autores al considerar lo que afecta o no los requisitos (Bureau y Encarnação, 2006; Bureau, 2008). Debido a que estos supuestos se contradicen entre sí, una misma dieta puede considerarse deficiente o no según el modo de expresión del requerimiento de EAA. De manera similar, la elección de la variable de respuesta probablemente influya en las estimaciones de los requisitos, ya que introduce otro nivel de variabilidad entre los experimentos. Las respuestas comúnmente utilizadas en este contexto incluyen expresiones de ganancia de peso (ganancia de peso absoluta, tasa de crecimiento específica, aumento porcentual, coeficiente de crecimiento de unidades térmicas, etc.), eficiencia alimenticia, retención de nitrógeno o, más raramente, concentración de EAA en sangre. La mayoría de los estudios consideran el crecimiento como la respuesta principal, pero no todos utilizan el mismo peso inicial en una especie determinada. Esto también es problemático, ya que los procesos biológicos que rigen el crecimiento de los peces son bastante dinámicos y cambian significativamente durante el ciclo de producción (Dumas et al. 2007), y es probable que los requisitos de EAA también cambien. Los modelos matemáticos completan el tríptico del aspecto analítico de la estimación de requerimientos de nutrientes. Se han utilizado diferentes modelos para describir la respuesta biológica de los peces, que normalmente sigue la ley del “rendimiento decreciente”. El modelo de línea quebrada (BLM) se ha utilizado mucho durante las últimas dos décadas. Sin embargo, cuando se compara con otros modelos no lineales, el BLM arroja estimaciones más bajas de manera consistente (Hernandez-Llamas, 2009). Por el contrario, Shearer (2000) señaló que los modelos no lineales podrían generar estimaciones de requisitos de hasta cinco veces los requisitos publicados.
Claramente, existe una necesidad imperiosa de revisar los requisitos de EAA para peces. Por lo tanto, el presente estudio primero revisó extensamente la literatura existente sobre los requisitos de EAA en peces y estandarizó sistemáticamente los datos relevantes. Luego, el conjunto de datos resultante se analizó utilizando varios modos de expresión, respuestas y modelos matemáticos para volver a evaluar el requisito de EAA en estas especies de peces comercialmente importantes.
2 Métodos y cálculos
2.1 Búsqueda de la literatura y criterios de selección
Se realizó una búsqueda exhaustiva de la literatura sobre los requisitos de la EAA utilizando bases de datos en línea, revistas revisadas por pares y libros. En este punto, el único criterio de selección fue mantener solo estudios sobre especies comercialmente relevantes (por ejemplo, salmonidae, cychlidae, cyprinidae), lo que resultó en 286 artículos. Hubo entre estos una gran variedad de objetivos, diseños experimentales y metodologías analíticas empleadas. Por lo tanto, se aplicaron criterios de selección adicionales al conjunto de datos para eliminar los estudios inadecuados para el presente metanálisis.
Dado que el crecimiento fue, con mucho, el tipo de respuesta más estudiado, solo se mantuvieron los estudios basados en ensayos de crecimiento en respuesta a niveles dietéticos graduados de un AA único. Por lo tanto, no se incluyeron experimentos basados en la medición de los niveles de AA en plasma o en la teoría de la proteína ideal (37 estudios de los 286 iniciales).
Otros criterios de selección fueron los siguientes: las prácticas de cría deben ser adecuadas para las especies de interés, incluida la calidad del agua. Se debe proporcionar información sobre la temperatura y la duración del experimento. Todas las dietas experimentales deben caracterizarse adecuadamente y cumplir con todos los demás requisitos nutricionales conocidos. Los estudios deben utilizar cinco o más dietas experimentales para garantizar la precisión de los análisis de regresión no lineal. Los datos, especialmente la composición de la dieta, deben informarse sobre la base de la materia seca, o se debe proporcionar suficiente información para calcular la composición de la materia seca de las dietas. No se incluyeron los estudios en los que la mortalidad fue alta y no se correlacionó con los niveles graduados de EAA. Finalmente, se debe informar el peso corporal individual inicial y final, así como el consumo de alimento, o se debe proporcionar información suficiente para calcularlos.
2.2 Combinaciones de variables y modelos
El metaanálisis se realizó como una matriz de 3x2x4: tres variables de factor (porcentaje de EAA de interés en la dieta seca, cantidad de EAA por MJ de energía digerible y cantidad de EAA de interés ingerido durante el experimento), y dos variables de respuesta (ganancia de peso por kg de peso corporal metabólico y coeficiente de crecimiento de la unidad térmica, TGC). Juntos, forman seis pares de variables. Finalmente, se utilizaron cuatro modelos de regresión (línea quebrada, cuadrático, cuadrático roto y cinético de saturación) para estimar los requisitos de EAA para cada par de variables y para cada estudio seleccionado.
2.2.1 Factor calculado y variables de respuesta
Aumento de peso, sobre la base del peso corporal metabólico (WGMBW, en g/kgMBW/d)

Coeficiente de crecimiento de la unidad térmica (TGC, adimensional)
Nivel dietético de AA a energía digestible (AADE, en g/MJDE)
AA ingeridos (AAI, en g)
Donde IBW y FBW son el peso corporal inicial y final individual (en g), respectivamente; d es la duración del experimento (en días), t es la temperatura promedio durante el experimento (en ⁰C); AAD es el contenido de la dieta del aminoácido de interés (% de materia seca); GED es el contenido energético bruto de la dieta (en MJ.kg⁻¹); ADCGE es el coeficiente de digestibilidad aparente de la energía bruta (%); FI es el consumo de alimento durante el experimento (en g, base de materia seca).
2.2.2 Modelos
El modelo de línea quebrada (BLM) se define de la siguiente manera:
Donde Y y X son la respuesta y las variables independientes, respectivamente; Xbp es la abscisa del punto de ruptura, definiendo así el nivel de requerimiento de AA; b1 y b2 son las intersecciones, y a1 y a2 las pendientes, de las 2 rectas, respectivamente. Tenga en cuenta que a2 no estaba restringido a cero.
El modelo cuadrático (QM) se define de la siguiente manera:
Donde Y y X son la respuesta y las variables independientes, respectivamente, mientras que a, b y c son parámetros de la ecuación. El requerimiento de aminoácidos es la abscisa de la raíz más pequeña que resuelve la ecuación de la parábola para Y=95% de la respuesta máxima.
La siguiente ecuación define el modelo cuadrático roto (BQM):

Donde Y y X son la respuesta y las variables independientes, respectivamente, y a, b y c son parámetros de la ecuación. Xjp es la abscisa del punto de unión, definiendo así el nivel de requisito de AA. Tenga en cuenta que dado que Xjp es una constante, la curva se convierte en una línea de pendiente cero para los niveles de aminoácidos por encima del requerimiento estimado.
Finalmente, el modelo cinético de saturación (SKM) se define de la siguiente manera:
Donde Y y X son la respuesta y las variables independientes, respectivamente; a es la asíntota de la respuesta máxima; b es la intersección; c es un coeficiente; d es un orden cinético. El requerimiento de aminoácidos se define como la abscisa donde la variable de respuesta alcanza el 95 % de su máximo previsto (es decir, el 95 % de a).
Las estimaciones de los requisitos de EAA se calcularon para cada una de las parejas de variables independientes/dependientes y para cada uno de los 4 modelos. Los resultados se clasificaron en las siguientes categorías: "sin ajuste" indica que el modelo no logró converger y, por lo tanto, no se generó una estimación de los requisitos. “Dentro de” significa que el modelo convergió y el requisito estimado cayó dentro del rango de niveles experimentales de EAA probados en un estudio determinado.
"Fuera" significa que el modelo convergió con éxito, pero el requisito estimado quedó fuera del rango de EAA de las dietas experimentales. “Sin ruptura”, para BLM y BQM, indica que solo se ajustó una de las dos curvas calculadas por el modelo, por lo que no se pudo determinar ningún punto de ruptura. Finalmente, los modelos de base cuadrática (BQM y QM) a veces convergían a una a positiva, lo que provocaba que el modelo tuviera un mínimo en lugar de un máximo. Este comportamiento se anotó como “mínimo”.
2.3 Análisis estadístico
Los modelos se programaron y ejecutaron utilizando SAS 9.4 (SAS Institute). Las cualidades de ajuste y el coeficiente de variación de las estimaciones de requisitos se probaron mediante pruebas logísticas y ANOVA de dos vías (pares de variables, modelos), respectivamente.
3. Resultados y discusión
3.1 Resultados del cribado
Durante la búsqueda bibliográfica se encontraron un total de 348 artículos revisados por pares que abarcan 52 años (1964 a 2016), de los cuales 48 no eran ensayos basados en el crecimiento, p. basado en formulaciones de proteínas ideales, o en cambios en los niveles plasmáticos de AA, artículos de revisión; por lo tanto, no se pudo incluir en el metanálisis. Sin embargo, otros criterios de selección rechazaron más de la mitad (52 %) de los 300 estudios restantes del metaanálisis (Figura 1). Las principales razones del rechazo incluyeron un crecimiento deficiente, falta de información (p. ej., datos de materia seca, consumo de alimento) o muy pocos niveles de clasificación (Tabla 1). El conjunto de datos final (Tabla 2) 145 estudios que cubrieron 37 especies de peces y 11 EAA. Alrededor del 72% de los estudios utilizaron 6 o 7 niveles graduados del EAA de interés. Un estudio utilizó 24 niveles graduados. Las variables más utilizadas para la estimación del requerimiento son el contenido de EAA de la dieta (% de materia seca) y la ganancia de peso (como SGR o % de PCI) como variable independiente y de respuesta, respectivamente. Los modelos más utilizados son el BLM y el QM.

Figura 1: Diagrama que ilustra la selección de estudios, desde el número total encontrado hasta el número de estudios adecuados que constituyeron nuestro conjunto de datos de trabajo
Tabla 1: Rechazo del estudio

El número de estudios excluidos es sorprendente. Destaca la calidad desigual del esfuerzo de investigación en esta área. También exacerbó la fragmentación del conjunto de datos final, tanto en términos de especies como de EAA. Por ejemplo, el 25 % de los estudios se centró en la trucha arcoíris, mientras que los estudios sobre otras especies, como la lubina, la platija japonesa o la tilapia del Nilo, representan cada uno el 3 % o menos del conjunto de datos.
De manera similar, la lisina es, con mucho, el EAA más estudiado (27 % de los estudios), mientras que solo cinco estudios (3 % del conjunto de datos, Figura 2) describen el requerimiento de fenilalanina. Además, estos cinco estudios se realizaron en cinco especies diferentes. Tal fragmentación constituye un gran desafío para la comprensión y definición de los requisitos de la EAA, y mucho menos para su racionalización.
3.2 Estimaciones de los requisitos de EAA
Tabla 2: Resumen de los estudios incluidos en el conjunto de datos final



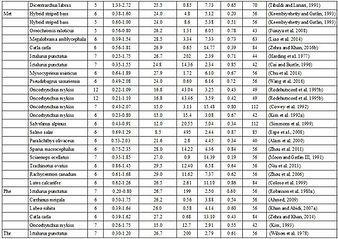

Figura 2: Representación de los once EAA en el conjunto de datos
En la Tabla 3 también vemos los requerimientos estimados con diferentes pares de variables. Si bien la influencia de las diferentes variables de respuesta se puede observar directamente, no tiene sentido comparar los requisitos calculados con diferentes modos de expresión, ya que se expresan en diferentes unidades.
En cambio, el coeficiente de variación (CV) de las estimaciones de requisitos proporciona una buena indicación de la consistencia de la estimación entre pares de variables. Independientemente de la especie y el EAA, los CV rondan el 22 % (la mediana es del 15 % en todo el conjunto de datos), que es comparable al 34 % encontrado por Hauler y Carter (2001),
pero mucho menor que el 100 %+ descrito por Shearer (2000). Bodín et al. (2009) encontraron que la elección de la variable de respuesta afecta el requerimiento relativo de lisina en la trucha arcoíris, pero no el requerimiento absoluto (es decir, el consumo de lisina). En nuestro conjunto de datos, las estimaciones de requisitos absolutos son numéricamente más cercanas entre las variables de respuesta que las estimaciones de requisitos relativos. Sin embargo, la variabilidad de las estimaciones nos impide establecer significación estadística. Al centrarse en los requisitos para la trucha arcoíris, muestra que ninguno de los cuatro modelos matemáticos da como resultado estimaciones más precisas. Asimismo, y contrariamente a las conclusiones de Hauler y Carter (2001) o Bodin et al. (2009), no observamos una diferencia significativa en la desviación estimada entre los dos modos de expresión relativos (% de materia seca de la dieta y g/MJDE) y absolutos (g ingeridos/kg MBW/d). Sin embargo, la relación entre el requerimiento de lisina en la trucha arcoíris y el PCI dependía de la unidad de requerimiento (Figura 3): hay una regresión lineal significativa cuando el requerimiento se expresa en g/kg MBW/d (P=0.0041, R²=0.482) , pero no cuando se expresa como porcentaje de materia seca de la dieta (P=0.3170, R²=0.077). Por lo tanto, las diferencias en el tamaño de los peces explican parte de la desviación observada y se pueden cuantificar mejor utilizando un modo de expresión absoluto basado en la ingesta.

Figura 3: Relación entre el peso corporal inicial (IBW, g) y el requerimiento de lisina de O. Mykiss expresado como porcentaje de materia seca de la dieta o como cantidad ingerida.
En general, la variabilidad en las estimaciones de requisitos limita nuestra interpretación de los requisitos de EAA en peces. También subraya la necesidad de identificar mejor los factores que rigen los requisitos de EAA. Esto se refleja claramente en el debate sobre el modo de expresión del requisito de EAA. Los tres modos de expresión elegidos para este metaanálisis representan tres enfoques fundamentalmente diferentes que se basan en supuestos mutuamente excluyentes.
Por lo tanto, existe una gran necesidad de investigar más a fondo los determinantes de los requisitos de EAA y, en particular, su dependencia de los ingredientes dietéticos o la ingesta de alimentos.
3.3 Limitaciones generales en el conjunto de datos
Una de las piezas de información más grandes que faltan en el conjunto de datos actual es la digestibilidad. De hecho, solo 15 de los 145 estudios seleccionados midieron e informaron la digestibilidad de las proteínas, y ninguno midió la digestibilidad de los EAA individuales. Además, se formularon dietas experimentales utilizando una amplia gama de ingredientes.
En los estudios seleccionados, las dietas se formularon normalmente utilizando una mezcla de proteínas naturales (harina de pescado, proteínas de soja y maíz, etc.) y aminoácidos cristalinos. No solo el EAA proporcionado por estos ingredientes probablemente tenga una digestibilidad diferente, sino que también puede haber interacciones relevantes entre los aminoácidos y con otros componentes no proteicos.

Figura 4: Respuesta de crecimiento del rodaballo (Psetta maxima) a niveles graduados de arginina en la dieta, que ilustra un patrón lineal. Adaptado de Fournier et al. (2003).
Estas interacciones pueden afectar la disponibilidad de EAA para el animal, de ahí el requisito aparente de EAA. Por ejemplo, se sabe que el fitato se une a los aminoácidos, especialmente a los aminoácidos básicos como la lisina, la histidina y la arginina, lo que limita su biodisponibilidad para el animal (Chowdhury et al., 2012; Selle et al., 2006; Storebakken et al. , 2000). Por lo tanto, los datos de los estudios que utilizan ingredientes ricos en fitato, como las proteínas vegetales, deben interpretarse con cuidado
Tabla 4: Comparaciones de las estimaciones del requerimiento de lisina en la trucha arcoíris O. mykiss, según la elección de variables y el modelo matemático

Tabla 3: Estimaciones de los requisitos de aminoácidos esenciales calculados utilizando el modelo cuadrático (QM)

Los valores son estimaciones del requerimiento en la unidad indicada, ± el coeficiente de variación
El peso corporal inicial de los peces también varía mucho entre los estudios, de 0,16 g a 662 g, aunque el 60 % de los estudios comenzaron con peces de entre 0,5 y 25 g. Sin embargo, se sabe que la utilización de nutrientes cambia con el tamaño de los peces, incluso en las primeras etapas juveniles (Dumas et al. 2007). Por lo tanto, considerar los requisitos de EAA en la etapa de crecimiento probablemente introduce una fuente de variación en las estimaciones de requisitos.
Finalmente, solo una minoría de los estudios informó sobre el análisis proximal final del pescado o los resultados de la deposición de proteínas. Esto nos impidió incluir variables como la retención de proteínas en nuestro análisis, que también se han debatido como una forma posible de expresar los requisitos de EAA.
3.4 Elección del modelo matemático y diseño experimental
Aunque la elección del modelo matemático no mejora la variabilidad de las estimaciones de requisitos, hubo diferencias significativas en el rendimiento y la calidad del ajuste. BLM, BQM y QM siempre convergieron, mientras que SKM no lo hizo en el 19,9% de los estudios seleccionados, todos los pares de variables combinados. Cabe señalar aquí que un modelo convergente no es suficiente para producir una estimación exacta y/o precisa del requisito. La Tabla 5 muestra a qué parámetros de diseño experimental son más sensibles los modelos y la calidad del resultado más allá de la convergencia del modelo. Tanto BLM como SKM son sensibles al número de dietas experimentales. Por ejemplo, cada dieta adicional en el diseño experimental hará que la BLM tenga 0,23 veces más probabilidades de producir un mínimo en lugar de un máximo (o 4,35 veces más probabilidades de producir un máximo, p=0,0028). El modelo SKM, aunque más cercano a la realidad biológica, exige un cuidadoso diseño experimental. Aunque un bajo número de dietas no aumentará significativamente el riesgo de ajuste fallido, cada dieta adicional hará que la SKM sea 0,89 veces más probable de dar como resultado una estimación fuera del rango experimental (p=0,0059). Esto es problemático y un ajuste "externo" no debe considerarse preciso.
Aunque el crecimiento de los peces se utilizó como criterio de selección para constituir el conjunto de datos de trabajo, la relación maxFBW/IBW aún afectó significativamente la forma en que los modelos QM y BQM ajustaron los datos (Tabla 5). El alto aumento de peso en los mejores (presumiblemente completos) tratamientos durante el experimento dio como resultado un riesgo reducido de producir una estimación fuera del rango experimental: razón de probabilidad de 0,63 (p = 0,0111) y 0,71 (p = 0,0084) para BQM y QM, respectivamente. En otras palabras, la ganancia de peso suficiente durante el experimento contribuye a las estimaciones dentro del rango experimental y, en ese sentido, contribuye a la precisión de las estimaciones. Más sorprendentemente, el BQM también tiene una tendencia significativa a ajustarse a un mínimo (es decir, pendiente negativa del segmento cuadrático) en valores altos de maxFBW/IBW. Tras una inspección más detallada de tales estudios, la respuesta del crecimiento de los peces a los niveles graduados de EAA siempre es bastante lineal, como se ilustra en la Figura 4. Sin embargo, todos los modelos se basan en la ley de "rendimientos decrecientes" y, por lo tanto, son más adecuados para situaciones que ilustran tal patrón. En el 50% de los casos de BQM fallidos (y el 97% de los BLM fallidos), el modelo no pudo ajustarse a una de las 2 líneas. Esto destaca que los modelos son sensibles a la presencia tanto de la porción ascendente inicial de la curva como de la siguiente meseta. La sensibilidad de los modelos al primero está bien ilustrada por la relación entre el peso más bajo y más alto de los peces al final del estudio (minFBW/maxFBW).
El minFBW/maxFBW fue, con mucho, la métrica más impactante con respecto al rendimiento y la calidad del ajuste en todos los modelos. Si uno pudiera imaginar un aumento de 1 unidad en la relación minFBW/maxFBW, el BQM y el BLM tendrían 106,40 (p=0,0307) y 26,28 (p=0,0319) veces más probabilidades de fallar en encontrar un punto de ruptura, respectivamente. El BLM también tendría 83,75 veces más probabilidades de ajustarse a una primera línea con pendiente negativa (p=0,0392). Finalmente, el SKM sería 3,20 (p=0,0170) y 30,73 (p<0,0001) veces más probable de encontrar una estimación
fuera del rango experimental y fallar por completo, respectivamente. Evidentemente, los estudios que no logran maximizar la diferencia en los pesos promedio finales entre tratamientos al final del experimento corren un gran riesgo de no poder estimar con precisión el requerimiento. Esto ocurre cuando la dieta basal no es lo suficientemente deficiente y/o el contenido de aminoácidos en otras dietas experimentales no alcanza un nivel lo suficientemente alto como para producir una meseta clara. En consecuencia, se debe prestar especial atención a las formulaciones dietéticas para crear dicho patrón.
La discusión anterior destaca los factores que los nutricionistas deben considerar cuidadosamente al desarrollar el diseño experimental y formular dietas experimentales.
Tabla 5: Razón de probabilidades de maxFBW/IBW, minFBW/maxFBW y número de dietas que afectan el resultado de ajuste de los modelos de requisitos.

Nota: valora la razón de probabilidades de que ocurra un resultado frente a un ajuste convergente que da como resultado una estimación dentro del rango experimental. Mínimo: el modelo tiene un mínimo en lugar de un máximo; Sin punto de ruptura: no se pudo determinar el punto de ruptura; Ajuste fallido: el modelo no logró converger; Fuera de estimación: el requerimiento estimado está fuera del rango experimental. NS: razón de momios no significativa diferente de 1; Las celdas vacías indican que este resultado no es aplicable al modelo.

Figura 5: Respuesta del crecimiento del rodaballo (Psetta maxima) a niveles escalonados de arginina en la dieta, que ilustra un patrón lineal. Adaptado de Fournier et al. (2003).
Nuestros resultados muestran que ninguno de los cuatro modelos da como resultado una precisión significativamente mejorada de la estimación de requisitos. El BLM ha sido un modelo de elección durante décadas, debido a su simplicidad y facilidad de ajuste, especialmente en un momento en que las computadoras no eran tan omnipresentes. Sin embargo, se han puesto de manifiesto las limitaciones del BLM (Hernandez-Llamas, 2009) y desde entonces se han sugerido otros modelos. Sin embargo, no se puede suponer a priori que ningún modelo sea superior en todas las situaciones, y se debe implementar un cuidadoso procedimiento de selección de modelos para cada experimento como parte de la evaluación estadística de los resultados. Para ello existen varios métodos, aunque el método Akaike Information Criterion (AIC) (Akaike, 1974; Arnold, 2010), que equilibra la bondad de ajuste y la parsimonia de los modelos, es práctico y orienta al nutricionista en su elección de selección del modelo (Salze et al. 2017).
4 Conclusiones y recomendaciones
La estandarización del cuerpo actual de literatura científica, tal como se lleva a cabo actualmente, no condujo a estimaciones de requisitos más claras y precisas. Sin embargo, destacó algunos puntos importantes.
Primero, la calidad de los estudios publicados es muy variable: menos de la mitad de los estudios relevantes se consideraron adecuados para nuestro conjunto de datos final. Las principales razones de estos rechazos incluyen un crecimiento deficiente y la falta de información reportada, como el contenido de humedad de la dieta.
Las investigaciones futuras de los requisitos de EAA deben centrarse en especies que no sean salmónidos, diferentes etapas de vida y EAA menos estudiados, como taurina, fenilalanina, leucina o histidina, para remediar la fragmentación del conocimiento. Los ensayos deben usar no menos de 6 dietas experimentales, más si las instalaciones y los recursos disponibles lo permiten. Debe evitarse el uso del BLM, ya que ahora se ha demostrado claramente que subestima el requisito. La elección del modelo matemático debe seguir luego una evaluación racional de la calidad del ajuste. SKM, QM o BQM son todos modelos válidos, y ninguno debe asumirse a priori como inherentemente superior a otro. Además, es importante que el número y el rango de niveles dietéticos graduados de la prueba EAA se elijan de modo que los segmentos ascendentes y de meseta de la curva puedan establecerse claramente con el requerimiento aproximadamente en el medio.
Finalmente, la pérdida efectiva de conocimiento de TI por información no reportada es muy contraproducente.
Por lo tanto, es muy importante que los nutricionistas de peces acuerden un sistema estándar y sistemático para informar y capturar los resultados de dicho ensayo.