Carbohidratos
4.1 Carbohidratos
Después de las proteínas y lípidos, los carbohidratos representan el tercer grupo de compuestos orgánicos más abundantes en el cuerpo animal, En contraste, los carbohidratos constituyen los nutrientes orgánicos principales del tejido vegetal. El grupo de los carbohidratos incluye importantes compuestos como la glucosa, fructosa, sucrosa, almidón, glucógeno, quitina y celulosa.
Los carbohidratos son definidos como aquellas substancias que contienen carbono, hidrógeno y oxígeno, con los dos últimos elementos presentes en la misma proporción que en el agua (p. ej. Cx (H2O) y). Aunque esta definición es satisfactoria par la mayoría de los compuestos presentes dentro de este grupo, algunos carbohidratos contienen una proporción menor de oxígeno, que en el agua, o bien existen derivados de carbohidratos que pueden contener nitrógeno y azufre.
4.1.1 Clasificación
Acorde a su estructura química, los carbohidratos se pueden dividir en dos grupos principales, azúcares y no azúcares (Tabla 8). Los azúcares más simples se denominan monosacáridos, mismo que pueden dividirse en 5 subgrupos dependiendo del número de átomos de carbono presentes en la molécula:
Estos monosacáridos pueden unirse entre sí (con la eliminación de agua) para formar di, tri ó polisacáridos, conteniendo dos, tres, ó más unidades o residuos de monosacáridos. Por lo que los carbohidratos denominados “no azúcares”, son aquellos que contienen más de 10 unidades de monosacáridos y que no poseen un sabor dulce. Los no azúcares pueden dividirse en dos subgrupos, hemopolisacáridos y heteropolisacáridos; consistiendo los primeros en unidades de monosacáridos idénticas, y los últimos son mezclas de diferentes unidades de monosacáridos.
4.1.2 Monosacáridos
En general todos los monosacáridos son solubles en agua, escasamente en etanol e insolubles en éter, son activos ópticamente, poseen propiedades reductoras (p. ej. reducen la solución de Fehling) se representan con la fórmula general CxH2xOx, y generalmente son de sabor dulce.
La fórmula estructural de los monosacáridos más comunes se representa como sigue:

En las fórmulas estructurales representadas, la “D” representa la configuración dirección del grupo aldehído (OH) ubicado en el átomo de carbono próximo al último carbono del grupo aldehído. Por ejemplo, en el caso del D(+)gliceraldehído y D(+)-glucosa, el grupo hidroxilo del penúltimo átomo de carbono (p.ej. C2y C5 respectivamente) están del lado derecho en relación al grupo aldehído (RCHO) de la parte superior de la fórmula. De igual modo, el símbolo (+) ó (-) indica la dirección de la rotación óptica producida cuando una solución del azúcar se coloca bajo el campo de
un polarímetro; dextrógiro (en el sentido de las manecillas del reloj, -). Virtualmente todos los monosacáridos naturales son miembros de las series D: consecuentemente, la configuración en torno al penúltimo átomo de carbono es la misma que la del Dgliceraldehído.
Además, todos los monosacáridos existentes en la naturaleza, son dextrógiros, con excepción de la fructuosa y la eristrosa.
Existen evidencias suficientes para sugerir que los monosacáridos también pueden existir en las formas moleculares cíclicas o anillos. Por ejemplo, las dos formas cíclicas de la D-glucosa que existen en la naturaleza α Dglucosa y β-D-glucosa. como en la fórmula estructural, la diferencia entre estas dos formas cíclicas depende de la configuración o dirección del grupo hidroxilo en el átomo de carbono¹.

Tabla 8. Clasificación de carbohidratos.

La importancia biológica de la diferencia estructural entre la α y la β-Dglucosa debe ser señalada; la configuración estructural determina las propiedades físicas y
consecuentemente biológicas de los polisacáridos constituidos por unidades individuales de monosacáridos. Por ejemplo la celulosa, polisacárido compuesto por cadenas en zigzag de unidades de β-glucosa: mientras que el almidón y glicógeno, polisacáridos constituidos por cadenas helicoides ó ramificadas de α-glucosa, son más reactivos desde el punto de vista biológico.
En este momento se debe mencionar que los monosacáridos rara vez están directamente involucrados en reacciones bioquímicas intracelulares, sin embargo son primeramente transformados en algún derivado del monosacárido.
Derivados importantes de monosacáridos incluyen el éster de azúcar fosfato (D-glucosa-6-fosfato, D-glucosa-1fosfato, D-fructuosa-6-fosfato y diésteres de fosfato), azúcares amino (D-glucosamina), azúcares-ácido (ácido glucónico y ácido glucurónico) y azúcares-alcohol (sorbitol).

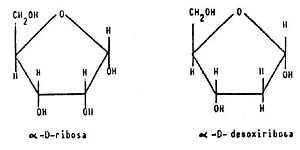
Pentosas
Importantes pentosas de monosacáridos incluyen la L-arabisona, D-xilosa, D-ribosa. Desde el punto de vista nutricional, la pentosa más importante es la D-ribosa y sus derivados D-desoxiribosa y ribitol. Por ejemplo, la D-ribosa y la D-desoxiribosa son componentes esenciales del ácido ribonucleico (ARN) y el ácido desoxirribonucleico (ADN), respectivamente y el ribitol es un componente esencial de la riboflavina.
Hexosas
Glucosa: Existe en su forma libre en tejidos de vegetales, frutas, miel y en la sangre. En la mayoría de los ingredientes alimenticios naturales, la glucosa existe en forma combinada, tanto con un monosacárido como un componente exclusivo de los disacáridos (p. ej. maltosa) y de polisacáridos (p. ej. almidón, glicógeno, celulosa) ó bien combinada con otros monosacáridos en forma de lactosa (azúcar de la leche), sucrosa y heteropolisacáridos. En el proceso de elaboración de la cerveza y vino, la glucosa es fermentada por la acción de levaduras, para producir alcohol y dióxido de carbono.
De igual modo, en el proceso de maduración de las frutas, el contenido de azúcar libre o glucosa aumenta, conforme disminuye el contenido del almidón.
Fructuosa: A semejanza de la glucosa, la fructuosa existe en su forma libre en los jugos de vegetales, frutas y en la miel. Es un componente del disacárido sucrosa y es el azúcar más dulce que existe en la naturaleza (p. ej. es responsable del sabor excepcionalmente dulce de la miel).
Galactosa: Aunque no existe en forma libre en la naturaleza, se presenta como un componente del disacárido lactosa y de muchos polisacáridos, incluyendo los galactolípidos, gomas y mucílagos.
4.1.3 Disacáridos
Los disacáridos están formados por dos azúcares hexosas, de cuya unión se elimina como residuo el agua:
Los disacáridos de mayor importancia que existen en la naturaleza son la maltosa, sucrosa y lactosa.
Maltosa: Está constituida por dos moléculas de glucosa unidas mediante un enlace α-1,4-glucosídico. La maltosa es un azúcar reductor, soluble al agua.
La maltosa no se encuentra en la naturaleza, pero es un producto obtenido durante la degradación enzimática del almidón. Por ejemplo, durante el proceso de germinación de la cebada, se obtiene maltosa a partir del almidón, gracias a la acción enzimática de la amilasa; una vez germinada y secada la cebada (que ahora se le denomina “malta”) se le emplea para la elaboración de cerveza y Whisky de malta.

Sucrosa: Esta constituida por una molécula de glucosa y una de fructosa unidas a través de un enlace α-1-β-2-glucosídico. Dado que los dos grupos reductores funcionales están involucrados en el enlace glucosídico, la sucrosa no posee propiedades reductoras.
La sucrosa está ampliamente distribuida en la naturaleza, se encuentra en la mayoría de las plantas; entre las fuentes ricas en sucrosa se incluyen al azúcar de caña (20% de sucrosa), azúcar de remolacha (15–20%) y zanahorias, La sucrosa es la azúcar utilizada a nivel doméstico para endulzar alimentos en casa.
Cuando la sucrosa es calentada a 160°C, forma azúcar de alfeñique (azúcar de cebada) y a 200°C forma caramelo. La remolacha y melaza de caña son productos secundarios de la agricultura, obtenidos mediante el proceso de elaboración se sucrosa a partir del azúcar de remolacha y de la caña de azúcar, respectivamente.

La lactosa o azúcar de la leche, es el principal azúcar presente en la leche y es exclusivo de los mamíferos. Forma aproximadamente el 40% del total de sólidos en la leche; el contenido total de lactosa en la leche de vaca y en la leche humana es de 4.6– 4.8% y 7%, respectivamente. La lactosa fácilmente sufre fermentación bacteriana, por ejemplo agriamiento de la leche por Streptococcus lactis, causado por la fermentación de lactosa a ácido láctico. A semejanza de la sucrosa, si la lactosa es calentada a una temperatura de 175° C forma lactocaramelo.
Las melazas son líquidos viscosos (20–30% de humedad), de color oscuro, de los que no se puede extraer más sucrosa mediante procesos de cristalización, debido a la presencia de cantidades apreciables de azúcares reductores (p. ej. glucosa) e impurezas.
Lactosa: esta compuesta de una molécula de glucosa y galactosa, unidas por un enlace β-1, 4-glucosídico. A semejanza de la maltosa tiene propiedades reductoras.

4.1.4. Homopolisacáridos
Estos carbohidratos son muy diferentes de los azúcares. Tienen un alto peso molecular y están compuestos de un gran número de hexosas o en el menor grado de residuos de pentosas. Muchos de ellos se les encuentra en vegetales y animales como material alimenticio de reserva (p. ej. almidón o glicógeno) o como elementos estructurales (p. ej. celulosa o quitina).
Almidón: esta compuesto de dos componentes estructurales, amilasa y amilopectina. Aunque las proporciones relativas entre la amilosa y la amilopectina en los almidones presentes en vegetales varían, dependiendo de las especies (20–30% de amilosa y 70–80% de amilopectina), la unidad fundamental de esos dos componentes estructurales es la alfa-D-glucosa. Por ejemplo, la amilasa consiste de largas cadenas no ramificadas, de 100 o más unidades de D-glucosa unidas entre sí por enlaces alfa-1, 4. De otra parte, la amilo pectina esta constituida por cadenas de unidades de D-glucosa altamente ramificadas (20–30 unidades por ramificación); los enlaces entre las unidades constitutivas son de tipo alfa-1, 4 y alfa-1, 6 (los enlaces alfa-1, 6 glucosídicos, son usados únicamente al inicio de las cadenas laterales).
El almidón es la forma química de almacenaje de azúcar o glucosa en los vegetales, se le encuentra en tallos, frutos, semillas y hojas, y representa la mayor reserva alimenticia de carbohidratos para los vegetales y consecuentemente constituye el mayor componente de carbohidratos en los alimentos de animales. Por ejemplo, el almidón puede representar hasta el 70% en peso de las semillas y hasta el 30% de los frutos, tubérculos o raíces. El almidón es almacenado dentro de los vegetales en forma de gránulos, cuya forma y tamaño varía de especie a especie. Cada gránulo esta rodeado por una capa delgada de celulosa que los hace insolubles al agua e indigestibles para los organismos no rumiantes, incluyendo peces y camarones, al ser ofrecidos en forma cruda o no cocidos. Al cocinarlos, por calentamiento en presencia de humedad, facilitará la ruptura de la membrana celulósica, dando lugar a la absorción del agua por el almidón, que en presencia de calor provoca la gelatinización del mismo, formándose una solución gelatinosa o pastosa. Cuando el almidón es expuesto a un calor seco, se formará dextrina, producto intermedio en la secuencia de su degradación: almidón→dextrina→maltosa→glucosa. Por ejemplo, el almidón presente en el pan, es convertido a dextrina cuando lo tostamos, la dextrina le da ese sabor característico al pan tostado. Glicógeno: Está compuesto por cadenas ramificadas de unidades alfa-Dglucosa, ligadas entre sí por enlaces alfa-1, 4 y alfa-1, 6; siendo los últimos los más abundantes en el glicógeno (en comparación con la amilopectina) debido a la presencia de ramificaciones más cortas y en mayor número, consistentes en 10–20 unidades de glucosa. El glicógeno es la forma en que los carbohidratos son almacenados en el cuerpo de los animales; en particular en músculo e hígado.
Celulosa: Está formada por cadenas muy largas de unidades de D-glucosa, enlazadas entre sí por uniones β - 1, 4, es un polisacárido muy estable y además es el carbohidrato más abundante en la naturaleza, siendo la estructura fundamental de la pared celular vegetal. La celulosa tiene una gran resistencia a la tensión y al ataque químico. Aunque la celulosa puede ser hidrolizada bajo un tratamiento con un ácido fuerte; con excepción de los microorganismos, pocos animales (excluyendo a los rumiantes) tienen las enzimas endógenas necesarias (p. ej. celulosa) capaces de hidrolizar y digerir la celulosa. Por ejemplo, las enzimas celulasas capaces de atacar la celulosa, únicamente se encuentran en semillas en germinación, hongos y bacterias (p. ej. las existentes en el tracto digestivo de los rumiantes). Un ejemplo de celulosa en una forma casi pura es el algodón.
Quitina: Está constituida de unidades repetidas de N-acetil-C-glucosamina, unidas por enlaces β-1, 4 y consecuentemente su estructura es similar a la celulosa.
La quitina es el principal componente estructural de la cutícula de los insectos y del esqueleto de crustáceos.

4.1.5 Heteropolisacáridos
En contraste con los homopolisacáridos, los heterosacáridos consisten en mezclas de diferentes unidades de monosacáridos y tienen un alto peso molecular.
Hemicelulosa: Esta compuesta principalmente por unidades de xilosa, unidas mediante enlaces β-1,4, pero también puede contener hexosas y azúcares ácidos (p. ej. ácido urónico). Estos polisacáridos normalmente acompañan a la celulosa en hojas, partes leñosas y semillas de vegetales superiores.
Son insolubles al agua y a semejanza de la celulosa no son fácilmente digeridas por otros animales que los rumiantes.
Gomas: Se les encuentra en la heridas de los vegetales y son compuestos muy complejos, al ser hidrolizados producen una gran variedad de monosacáridos y azúcares ácidos. Un ejemplo es la goma arábiga (goma de acacia).
Mucílagos: Son carbohidratos complejos presentes en ciertas plantas y semillas. Muchas algas, especialmente las marinas producen mucílagos, mismos que son solubles al agua caliente y forman un gel al enfriarse. El agar, un polímero de la galactosa con el ácido sulfúrico, es un mucílago o gel ampliamente utilizado, que se obtiene del alga marina roja (familia Gelidium). Otros ejemplos incluyen al ácido algínico, derivado de las algas cafés (familia Laminaria).
Sustancias pépticas: Son carbohidratos complejos que contienen ácido Dgalacto-urónico como principal constituyente. Naturalmente se encuentran en la pared celular primaria y en las capas intercelulares de vegetales terrestres, son particularmente abundantes en frutas de cítricos, azúcar de remolacha, manzanas y en algunas raíces de vegetales (p. ej. nabo). Como con los mucílagos, los ácidos pécticos muestran fuertes propiedades para la formación de gel y se les utiliza en la preparación de mermeladas.
Mucopolisacáridos: Son carbohidratos complejos que contienen azúcares amino y ácido urónico y constituyen las secresiones mucosas de los animales. Son de naturaleza ácida y pueden ser ricos en grupos éster-sulfato, mucopolisacáridos importantes incluyen al sulfato de condroitina (presente en el cartílago, hueso, válvulas cardiacas, tendones y en la cornea del ojo) heparina (anticoagulante presente en vasos sanguíneos, hígado, pulmones y bazo) y el ácido hialurónico (lubricante viscoso presente en piel, humor vítreo del ojo, líquido sinovial de articulaciones y el cordón umbilical en mamíferos).
Finalmente los mucopolisacáridos constituyen el componente principal de la pared celular de muchas bacterias; el mucopolisacárido de la pared celular consiste en unidades alternantes de ácido N-acetilmurámico y N-acetil-glucosamina, unida a pequeñas cadenas de peptidos.
4.2 Función de los carbohidratos
Los carbohidratos son sintetizados por todos los vegetales verdes, a través del proceso denominado fotosíntesis, que se representa como sigue:
Tanto en el hombre como en los animales terrestres, los carbohidratos suministrados en la dieta son la principal fuente de energía metabólica (ATP). Esta reacción se representa de la siguiente manera:
¹ ATP. ó Trifosfato de Adenosina, contiene enlaces de “alta energía”, y representa la manera en la cual una gran parte de la energía liberada es almacenada dentro del cuerpo
En peces y camarones no se ha establecido un requerimiento absoluto de carbohidratos en la dieta.
Esto contrasta marcadamente con lo establecido para las proteínas y lípidos, nutrientes para los cuales ya se han establecido requerimientos dietéticos específicos para ciertos aminoácidos y ácidos grasos esenciales. En gran medida esto se debe a:
-
Los hábitos alimenticios carnívoros/omnívoros de la mayoría de las especies d peces y crustáceos cultivados.
-
La habilidad de los peces y camarones para sintetizar carbohidratos (p. ej. glucosa) a partir de substratos que no sean carbohidratos, tales como proteínas y lípidos (proceso denominado gluconeogénesis).
-
La habilidad de los peces y crustáceos para satisfacer sus requerimientos energéticos a partir del catabolismo únicamente de proteínas y lípidos, si es necesario.
Sin embargo, a pesar de la aparente ausencia de un requerimiento dietético de carbohidratos para peces y crustáceos, no existe duda que los carbohidratos realizan importantes funciones biológicas en el cuerpo del animal. Por ejemplo, la glucosa producto final de la digestión de carbohidratos en los animales, sirve como la principal fuente energética para el tejido nervioso y cerebro y como intermediario metabólico para la síntesis de muchos compuestos biológicamente importantes, incluyendo el exoesqueleto quitinoso de los crustáceos, los ácidos nucleicos ARN y ADN, y los mucopolisacaridos de las secreciones mucosas.
A pesar de que los carbohidratos pueden ser considerados nutrientes no esenciales en la dieta de peces y crustáceos, su inclusión en las dietas de engorda está garantizada debido a:
-
Representan una fuente económica de energía dietética muy valiosa para aquellas especies de peces y crustáceos no carnívoras.
-
Su uso cuidadoso en dietas para engorda puede representar un ahorro en referente a la utilización de la proteína, insumo más valioso para el crecimiento en lugar de fuente de aprovisionamiento energético (procedimiento denominado “sustituto proteínico”).
-
Al ser empleados como ligantes sirven como constituyentes dietéticos esenciales, para la elaboración de dietas estables en el agua (por ejemplo: almidón gelatinizado, alginatos, gomas).
-
Ciertas fuentes de carbohidratos sirven como constituyentes dietéticos que aumentan la palatabilidad del alimento y disminuyen el contenido de polvo el alimento terminado (p. ej. melaza de caña o remolacha).
4.3 Metabolismo de los carbohidratos
Aunque el glicógeno constituye la principal fuente energética para el metabolismo anaeróbico (glicólisis) en el músculo blanco del pez durante el “rompimiento” en el nado, la habilidad del hígado y tejidos para almacenar glicógeno es limitada; los carbohidratos totales, en forma de glicógeno constituyen menos del 1 % del tejido corporal húmedo (Cowey & Sargent, 1979). En contraste, se ha visto que los juveniles del camarón P. japonicus, contienen glucosamina y trehalosa, como principales formas de carbohidratos en su tejido corporal (Kanazawa, 1983).
Al contrario de los mamíferos omnívoros, los peces no movilizan rápidamente el glicógeno del hígado cuando son mantenidos en ayuno. De hecho se ha detectado que durante el ayuno de los peces, la oxidación de substratos diferentes a carbohidratos tiene prioridad sobre la movilización e hidrólisis del glicógeno. Lo cual sugiere que la capacidad de los peces para oxidar anaeróbicamente la glucosa es limitada. Es por ello que la gluconeogénesis puede tener un papel fundamental en el mantenimiento de los niveles de azúcar en la sangre de peces hambrientos o en ayuno. Por ejemplo, para peces cultivados que se han alimentado con una dieta de elevado contenido proteínico, es probable que la demanda energética de los tejidos (p. ej. cerebro y tejido nervioso) que catabolizan glucosa, sea cubierta a través de la gluconeogénesis (a partir de aminoácidos y del triacilgliserol) más bien que la glicogénolisis (para revisión consultar a Walton y Cowey, 1982). Sin embargo, estudios con anguila (A. japonica) parecen contradecir la hipótesis arriba señalada (Degani, Viola y Levanon, 1986).
4.4 Utilización de carbohidratos
La habilidad de peces carnívoros para hidrolizar o digerir carbohidratos complejos es limitada debido a la debilidad en la actividad amilótica en tracto digestivo (Spannhof y Plantikow, 1983). Así, para especies tales como la trucha, conforme aumenta la proporción de almidón en la dieta, disminuye su digestibilidad (Singh y Nose, 1967; Bergot y Breque, 1983). Aún más, en ensayos de alimentación de mayor duración con peces carnívoros (p. ej. salmónidos) se ha observado que elevados niveles de carbohidratos en la dieta, disminuyen el crecimiento, elevan los niveles de glicógeno en hígado y eventualmente causan mortalidad (Phillips et al., 1977). Por el contrario los peces omnívoros o herbívoros de agua caliente, tales como la carpa (C. carpio), bagre de canal (I. punctatus), tilapia (O. niloticus) y la anguila (A. japonica) han mostrado ser más tolerantes para niveles elevados de carbohidratos; siendo utilizados más eficientemente los carbohidratos como fuente de energía o bien el exceso es almacenado en forma de lípidos corporales (Chiou y Ogino, 1975; Robinson y Wilson, 1985; Adreson et al., 1984; Degani, Viola y Levanon, 1986).
También se ha visto que la utilización de carbohidratos ofrecidos en la dieta varía con la complejidad o estructura química de la fuente de carbohidratos usada (los polisacáridos y disacáridos digestibles tienen un mayor efecto benéfico en el crecimiento que los monosacáridos: peces - Pieper y Pfeffer, 1980; Robinson y Wilson, 1985; Anderson et al., 1984; camarones - Alava y Pascual, 1984; Deshimaru, 1981¹; Kanasawa, 1983) el estado físico de la fuente de carbohidratos utilizada (almidones cocidos o gelatinizados tienen una mayor disgestibilidad y efecto benéfico sobre el crecimiento, en comparación con los almidones naturales o crudos: peces - Spannhof y Plantikow, 1983; Bergot y Breque, 1983; Robinson y Lovell (1984) y el consumo de alimento diario (un régimen de alimentación restringido tiene un efecto benéfico sobre la digestibilidad de almidón: peces - Bergot y Breque, 1983). De la discusión precedente, parece que la habilidad de los peces o camarones para adaptarse a dietas con un elevado contenido de carbohidratos, dependen de su habilidad para convertir el excedente energético (p. ej. glucosa) en lípidos o aminoácidos no esenciales.
Dado que la mayoría de los peces cultivados tienen un tracto gastrointestinal corto, que no les permite desarrollar una flora bacteriana abundante (como en los rumiantes), la actividad intestinal de la celulosa, en peces, a partir de las bacterias residentes es muy débil o nula (Stickney y Shumway, 1974).
Consecuentemente la celulosa o “fibra cruda” (p. ej. carbohidratos de la dieta que son resistentes al tratamiento químico con ácidos o álcalis diluidos, incluyen a la celulosa y hemicelulosa) no tiene ningún valor energético para los peces, y un exceso en la dieta tiene un efecto deletereo sobre la eficiencia alimenticia y el crecimiento (Anderson et al., 1984; Poston, 1986; Hilton, Atkinson y Slinger, 1983; Bromley y Adkins, 1984).
¹ Furichi, Taira y Yone (1986): la disponibilidad de la glucosa en el cola amarilla (S. quinquerradiata) es más bajo que el alfa-almidón.